AGING IN OTHER SPECIES
“Discovery consists of looking at the same thing as everyone else and thinking something different.”
Albert Szent-gyorgyi
A foundational misconception of the aging paradigm is the presumption that biological aging is a unitary phenomenon that has the same root cause in all organisms. The Institute acknowledges that the primary symptom of biological aging – the accumulation of intrinsic damage causing physiological deterioration that results in progressive dysfunctionality of all organs and systems – is the same in all mammals. Where the position of the Institute differs from conventional wisdom is that it posits that the accumulation of damage in humans has a different root cause than the age-associated accumulation of damage in other mammals.
Defining Terms
One of the recurring themes of this website is that much of the confusion surrounding aging is the failure to develop clear definitions of critical terms. The use of the term “aging” to describe the physiological deterioration experienced by both humans and other mammals makes the effort to distinguish between the two traits almost impossible. To avoid this problem, the Institute introduces different terminology. When referring to the declines in physiological performance suffered by all mammals as they approach the end of life, this website uses the term Natural Progressive Decline (“NPD”). When referring to the progressive declines in physiological performance suffered by a typical human commencing relatively early in life, this website uses the term Functional Decline Syndrome (“FDS”).
“Lifespan” means the period of time that, on average, the members of a species actually live. “Longevity” is the period of time that a member of a species is expected to live in the absence of aging-independent causes of death such as predators, starvation and non age-associated disease. This website introduces the term “natural lifespan” of a species to mean the period of time before substantially all members of that species necessarily die from aging-independent causes of death in the evolutionary environment.
Animal Longevity
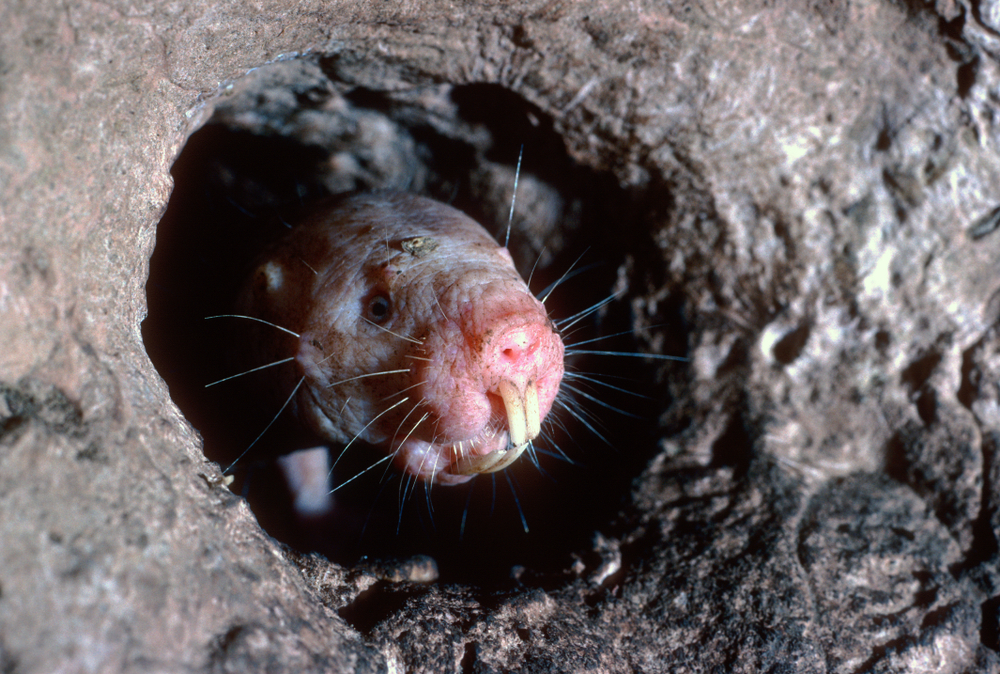
The naked mole rat can live for more than three decades.
Aging theorists have long noted the fact that the longevity of otherwise similar species can vary remarkably. An obvious example would be rodents. Even under ideal conditions, the lab rat will rarely survive for more than a few years; by contrast, the longevity of the naked mole rat is estimated to be more than three decades. All mammals (including humans) were descended from a common ancestor. The only explanation for the extreme variances in longevity is that longevity is genetically determined and developed through an evolutionary process in a way similar to the one that determines other species-specific traits.
How high can a bird fly? The answer varies dramatically depending on the species of bird. Where the ability to fly at great heights conferred a substantial evolutionary advantage, mutations that led to that result (at a cost to other traits) were passed down to subsequent generations. Each species of bird has different capabilities with respect to the ability to fly. All such capabilities are determined by the physiological makeup of the species, which is determined by natural selection.
PA proponents argue that the only physiological process that could have evolved to explain the variance in longevity among species would be their hypothetical aging mechanism. Per the PA argument, all organisms must possess complex evolved mechanisms that exist for the purpose of pro-actively limiting the organism’s lifespan beyond a species-specific age.¹
The Institute agrees with PA proponents that the only way to explain longevity variances among species is through the evolution of a physiological system that determines longevity. Where the Institute disagrees with the PA proponents is in identifying the maintenance system as being the critical physiological system. A core hypothesis derived from the New Paradigm is that the longevity of any species is a function of the capabilities of that species’ maintenance system. Like all other physiological systems, maintenance systems evolved in very different ways in different species.
In Their Evolutionary Environment, Animals Don’t Age
In their evolutionary environment, all mammals die from aging-independent causes of death before they show any signs of NPD.² NPD occurs in animals only because human interference allows them to outlive the effectiveness of their maintenance systems.
Substantially all domesticated animals have for many generations been the subject of human-designed breeding programs. As a result, in addition to not being exposed to the evolutionary environment, domesticated animals have, for generations, not been subject to the forces of natural selection.
Recent studies have found evidence of aging/progressive physiological deterioration in certain species in the wild. But the evolutionary environment is not the same as “the wild.” The effect of humans on the planet has been so pervasive that the current environment, even in the wild, is a far cry from the evolutionary environment. Human activities (e.g., hunting, pesticides such as DDT, etc.) have had a disproportionately adverse effect on predator populations. In the evolutionary environment, there would be a robust predator species for all other species. Animals tend to flock together not because they are going to protect one another (other than offspring), but rather to ensure that the least fit gets taken by predators.
The evolutionary environment is one in which each individual member of the species must acquire its own food and avoid/defend itself from a robust predator population. In an environment where predators are ubiquitous, and competing for sufficient food is a life or death challenge, symptoms of FDS – i.e., diminution in functionality – would likely prove to be fatal within a relatively short period of time. Human interference has altered the evolutionary environment by killing off a disproportionate number of predators, and domesticating and feeding many species. As a result, NPD appears to be a quite common phenomenon – domesticated animals typically outlive the natural lifespans of their species. Thus the conventional belief that NPD and FDS are the same phenomenon is not surprising.
NPD in Mammals Is Consistent with the Theory of Evolution
Distinguishing NPD from FDS
Conclusion
- Goldsmith TC, On the programmed/non-programmed aging controversy, Biochemistry (2012).
- Finch C.E., Longevity, Senescence and the Genome, Chicago University Press, (1990); Lack D, The Natural Regulation of Animal Numbers, Clarendon Press, (1954); Medawar P.B.,An Unsolved Problem of Biology, Lewis (1952). See also, Hayflick L, Entropy explains aging, genetic determinism explains longevity, and undefined terminology explains misunderstanding both. PLoS Genetics (2007).
This is the last essay in the section entitled “The Case for Paradigm Shift.” The next section is “The New Paradigm.”